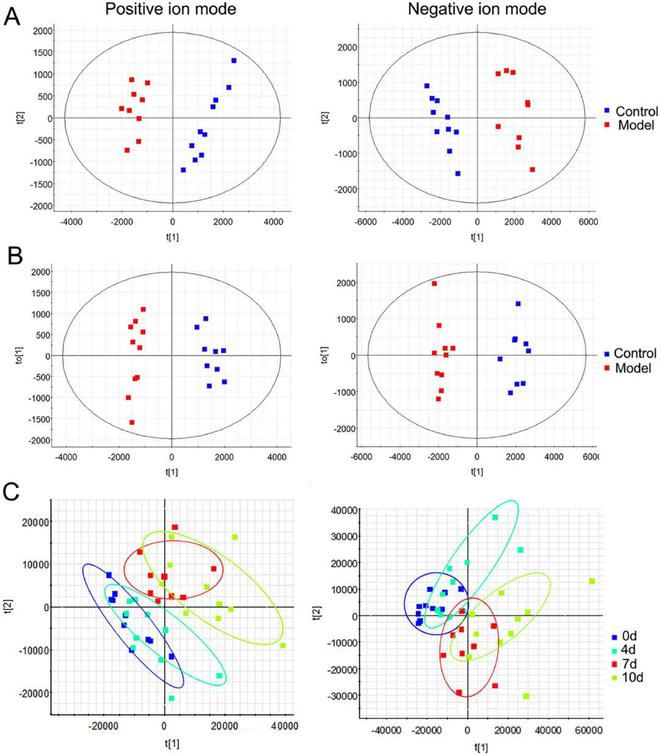
通过PCA和OPLS-DA的多元统计分析找出对照组和模型组之间的差异★■◆◆■,并评估PD对脂质代谢模式的影响★◆◆■■★。根据PCA评分图显示(图5A-B)■★◆★,对照组和模型组显著分为两组,表明两组内子宫组织脂质谱存在显著差异。OPLS-DA结果也表明对照组和模型组完全分离(图5C-D),正离子模式和负离子模式的R2Y和Q2值分别为0■■◆■◆.958、0■◆■★.928、0★◆★◆■.991和0.963■★◆。多元统计分析显示,该研究模型可靠且具有极佳的可预测性◆■◆★◆。
译名◆◆★:基于代谢组学和脂质组学探讨当归四逆汤改善原发性痛经脂质稳态的机制研究
结论:研究结果表明,DGSND通过调节脂质稳态对PD具有一种新的保护机制■■◆■★■。
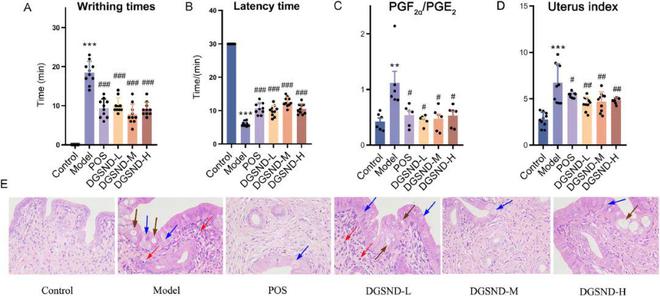
本研究采用扭动次数、潜伏时间、生化指标、子宫指标和组织病理学来评价PD动物模型和DGSND的治疗效果,其中扭动次数和潜伏时间是其重要指标。与对照组相比,PD大鼠在注射催产素后的潜伏时间约为6分钟■★◆■,且30分钟内的蠕动次数为18次,PGF2α/PGE2值显著升高(P0.01,图2A-C)。与模型组相比,经布洛芬或不同剂量DGSND治疗后,PGF2α/PGE2值显著下降(图2B,P 0.05)◆■★。模型组的子宫指数显著升高(图2D■◆◆★,P0.001)■★★◆,不同剂量的DGSND治疗后,子宫指数显著改善★★◆◆◆◆,与POS组相似(图2D,P0.001,P<0◆◆★★★★.05)。此外与对照组相比◆■★★★■,模型组子宫组织中出现了水肿、炎症浸润、空泡和疏松的细胞质,而POS组和DGSND组的水肿和炎症浸润较轻,空泡较少,细胞质相对致密(图2E)■◆■。以上结果表明◆■,DGSND可以有效缓解扭动时间、潜伏时间◆■■■、PGF2α/PGE2值、子宫指数和组织形态◆★■◆■,且与布洛芬组相比也显示出相似的治疗效果。
LIPEA还表明,坏死性凋亡和铁死亡与PD有关。脂质除了在能量储存和膜结构中发挥经典作用外,还是细胞增殖、分化和死亡的重要信号调节剂。越来越多的证据表明,甘油磷脂、鞘磷脂和脂肪酸在坏死性凋亡中起着至关重要的作用。其他一些子宫疾病的发生和发展也涉及到铁凋亡过程,而铁凋亡是一种依赖脂质的细胞死亡机制■◆◆■。PE(18:0/20★■◆■:4)和PE(18:0/22:4)是促进铁死亡的关键脂质,其表达可被DGSND显著逆转,提示PD的发生和发展可能涉及铁死亡,有待进一步研究。这也是本研究的局限性★★◆★■,鉴于不同类型脂质的功能,未来还需要进行精确★■、特异的靶向脂质组学分析◆★。
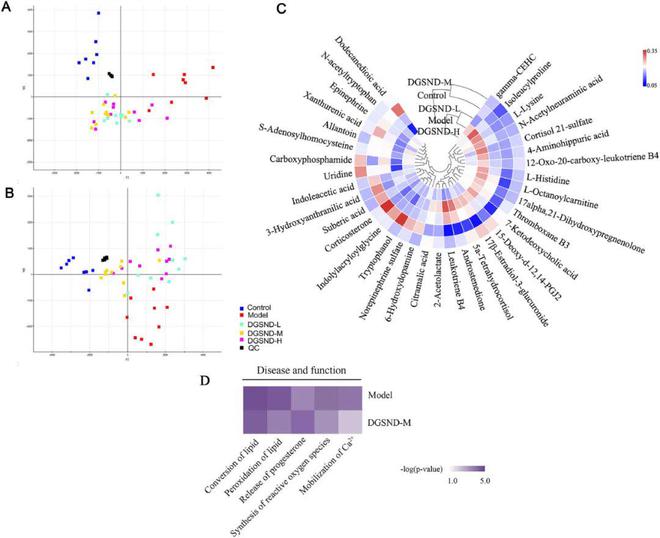
将重要的DGSND逆转生物标志物导入IPA平台,进一步用于挖掘其抗PD的机制(图4D)。疾病与功能模块的结果显示■◆■◆◆,与模型组相比◆★★,DGSND组的脂质转化、脂质过氧化、孕酮释放、活性氧合成和Ca2+的调动均显著逆转(图4D)。结合代谢途径分析,脂质代谢可能是DGSND治疗PD的潜在靶点★★◆■■◆。
S-腺苷同型半胱氨酸水平升高会影响蛋氨酸的胱硫醚代谢途径,最终影响牛磺酸的产生。牛磺酸可以抑制胆固醇的产生,调节抑制性神经递质的释放★★★■◆◆,从而减少类固醇激素的释放,缓解PD症状■■◆。
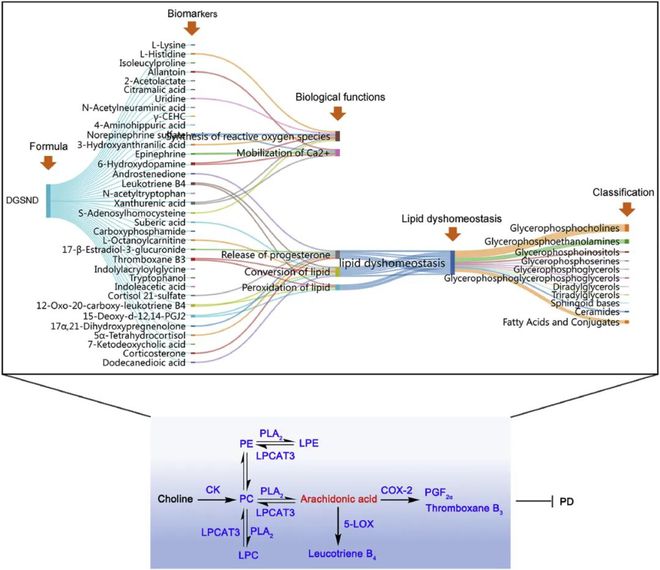
此外代谢组学研究还用于分析尿液样本的代谢特征★■★■■。PD引起的代谢改变是一个动态过程★★,共表征了35种与PD相关的生物标记物,其中类固醇激素代谢、氨基酸代谢和脂质代谢可受DGSND调控(图10)◆★。根据被调控生物标志物的生物学功能,构建了■◆★■“方剂-生物标志物-生物学功能-脂质稳态失衡-分类”的机制网络,以更好地理解药物效应(图10)★■◆★■◆。
目的:本研究致力于从代谢组学和脂质组学的角度探讨DGSND对PD的保护作用和潜在机制。
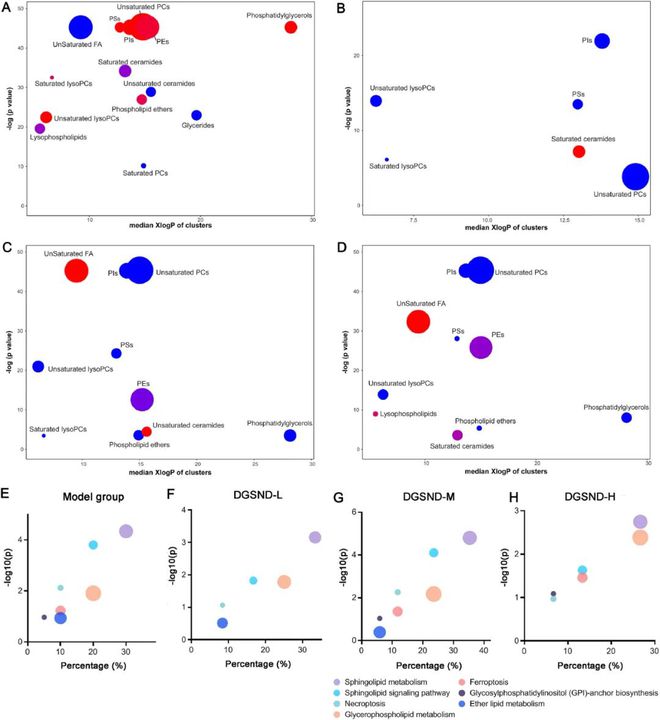
图7 受显著调控脂质的富集统计和LIPEA通路分析★★。富集统计分析■◆:(A)对照组与模型组■★■◆★■。(B) DGSND-L与模型组。(C) DGSND-M与模型组。(D) DGSND-H 与模型组。节点颜色显示化合物增加(红色)或减少(蓝色)的比例。紫色节点表示发生了增加和减少。节点大小代表每个聚类组中脂质的总数◆★★★。LIPEA通路分析:(E)对照组与模型组◆★。(F) DGSND-L与模型组★■◆■◆。(G) DGSND-M与模型组。(H) DGSND-H与模型组★■◆。
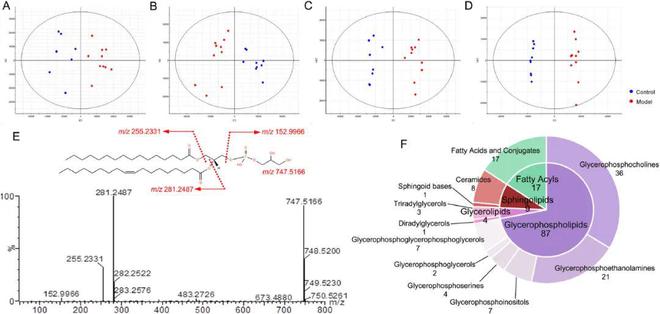
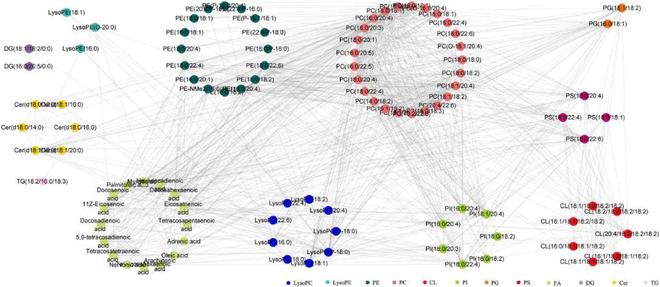
利用精确质量、MS/MS片段数据和在线数据库确定结构◆■★■,选择VIP1且P0.05的离子作为候选变量。在负离子模式下,保留时间为5.54 min的m/z 747◆■.5166离子被定性为潜在的生物标记物,其VIP为3.9。m/z 747■■■★■◆.5166的可能分子式为C40H77O10P◆★■◆★★。MS/MS质谱显示,m/z 747★★■■◆.5166被明确为m/z 281■■★.2487、255.2331,其中脂肪酰基在甘油主链的sn-1和sn-2位置酯化★◆◆■■■,m/z 152.9966为甘油磷酸酯极性头部基团(图5E)。该片段途径与数据库(HMDB)报告的磷脂酰甘油(16:0/18:1)相同★◆★◆■★,因此5★◆◆■.54_747■■◆.5166的脂质被初步鉴定为磷脂酰甘油(16◆◆■■:0/18◆■◆:1)。最后107种脂质被分为4个脂质类别和11个脂质亚类★◆◆★■★,包括9种溶血磷脂酰胆碱(lysoPCs)、27种磷脂酰胆碱(PCs)◆◆、4种溶血磷脂酰乙醇胺(lysoPEs)、17种磷脂酰乙醇胺(PEs)★★、7种磷脂酰肌醇(PIs)、4种磷脂酰丝氨酸(PSs)、2种磷脂酰甘油★■◆★◆◆、7种心磷脂(CL)、1种二酰甘油(DGs)★■、3种甘油三酯(TGs)、1种鞘氨醇(sphinganine)、8种神经酰胺(ceramides)、17种脂肪酸(FAs)(图5F■◆,表S8)。在本研究中最常见的脂类属于甘油磷脂■★■★★,占71.96%,脂肪酰基占15.89%,鞘脂占8★★.41%■★◆■★,甘油磷脂是次要成分,仅占3◆★■★.74%。
本研究平行处理了6份质控样品◆★★★,随机提取10个离子峰以覆盖不同的分析时间,从而考察尿液和子宫组织样品处理方法的稳定性。总离子色谱图见图S1-2。经统计分析,代表性离子峰面积的相对标准偏差(RSD)为0.9-5.5%(表S3-4)。在UPLC-MS分析过程中,为了验证系统的稳定性,分别对尿液和子宫组织质控样品进行了每10次进样分析,结果表明代表性离子的峰面积RSD为1.1-5■◆.3%(表S5-6)■■◆。结果表明尿液和子宫组织样品的分析具有良好的重复性和稳定性。
图S3显示了对照组和模型组尿样在正负离子模式下的色谱峰强度(BPI),显示了尿液代谢轮廓信息。PCA图显示了对照组和模型组样本显著聚类(图3A)◆★◆。OPLS-DA分析进一步确定了将对照组与模型组区分开来的变量(图3B)■★。OPLS-DA模型的参数R2Y和Q2在正离子模式下分别为0.993和0.968,在负离子模式下分别为0.965和0★◆.907■★★◆,表明本数据的拟合度和预测度符合要求。模型组在PD进展过程中的轨迹表明,PD的病理过程逐渐引起尿液代谢物的变化(图3C)★◆★。
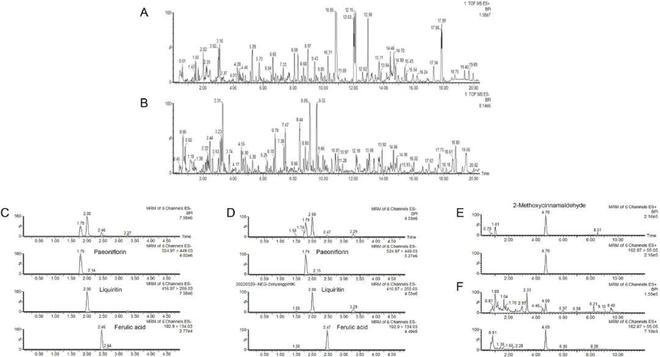
使用UPLC-Q-TOF/MS在正负离子模式测定DGSND的化学组成,离子色谱图如图1A-B所示,每种化合物的详细表征见表S1★◆。根据相关数据库初步鉴定了DGSND中的103种化合物,主要包括萜类、黄酮类、苯丙类等。通过 UPLC-TQD/MS 分析芍药苷■★◆、甘草苷、阿魏酸和2-甲氧基肉桂醛,评估了DGSND的质量(图1C-F),这些代表性成分在DGSND中的平均含量分别为1■★◆.675 mg/g、0.880 mg/g、0.196 mg/g 和 15★★◆■.594 mg/g(表S2)。
图2 DGSND对PD大鼠的影响。(A)注射催产素后的扭动次数■★。(B)扭动潜伏期。(C) PGF2α/PGE2值的表达水平。(D)子宫指数■★◆★■◆。(E) 子宫组织HE染色(放大×400)。蓝色箭头代表子宫内膜上皮细胞水肿,红色箭头代表炎症浸润,棕色箭头代表子宫内膜上皮细胞空泡。与对照组相比◆■,**P0.01,***P 0★■.001★■★;与模型组相比,#P0.05,##P0.01,###P0◆■■■★★.001■★。
本研究首次揭示了DGSND通过重塑脂质稳态对PD大鼠的新型保护机制。研究采用尿液代谢组学分析了PD大鼠的代谢特征★■◆★,功能分析揭示了脂质紊乱是DGSND治疗PD的主要靶点■◆◆。进一步的靶向脂质组学研究表明◆■★■★★,DGSND可以通过调节脂质相关信号通路和不同类别脂质之间的相互作用来改善PD疾病。
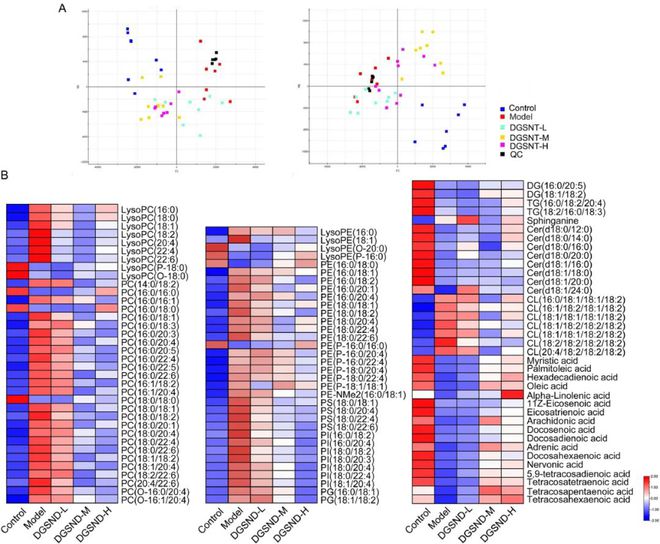
图6 DGSND干预后子宫组织代谢谱的多元统计分析。(A-B)正负离子模式下DGSND治疗PD的PCA评分图。(C)差异表达脂质的热图。
图3 尿液代谢轮廓和轨迹的多元统计分析■■★◆★。(A)对照组和模型组第10天代谢轮廓PCA评分图。(B)对照组和模型组第10天代谢轮廓的OPLS-DA评分图■◆。(C)模型组第0★★■、4、7和10天代谢轨迹的PCA评分图◆◆★★。
图1 DGSND的化学成分表征和质量控制■■★◆◆◆。(A-B)正负离子模式下DGSND的BPI色谱图■★★。(C)负离子模式下混合标准品(芍药苷、甘草苷和阿魏酸)和每种化合物的单独提取物的BPI色谱图。(D)负离子模式下DGSND样品和每种化合物单独提取物的BPI色谱图。(E)正离子模式下2-甲氧基肉桂醛和各提取物的BPI色谱图。(F)正离子模式下DGSND样品和2-甲氧基肉桂醛提取物的BPI色谱图。
【福利时刻】科研服务(点击查看):◆■★■◆◆、◆■、、、★◆■■、。咨询加微信:1278317307■◆。
本研究通过传统药理学、尿液代谢组学◆★■■■◆、功能分析和子宫组织脂质组学研究了DGSND对PD大鼠的保护作用。PD大鼠的代谢紊乱主要与类固醇激素代谢、氨基酸代谢和脂质代谢有关,DGSND治疗可改善这种失衡,尤其是脂质代谢。值得注意的是,不同脂质的具体功能仍有待进一步明确◆■◆。此外本研究加深了对PD病理生理机制的认识,为DGSND的保护作用提供了可用证据■★★★,同时也为中医机制研究提供了方法学参考■■。
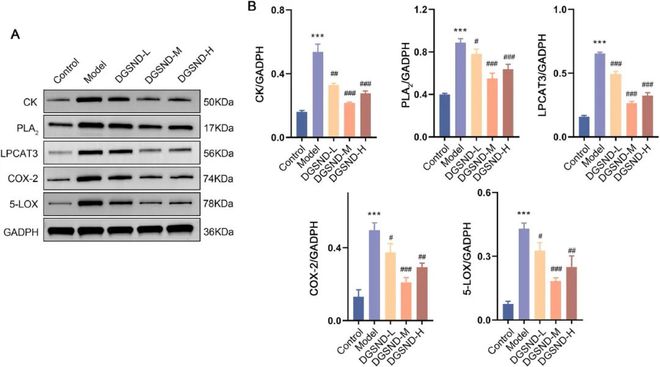
DGSND显著下调PD诱导的CK、PLA2、LPCAT3、COX-2和5-LOX过表达(图9)。上述酶在脂质代谢途径中至关重要,这也进一步表明DGSND可以改善脂质稳态以表现出改善PD的功效。
IPA分析结果表明,DGSND主要通过调节脂质平衡发挥抗PD作用(图4D)。PGs是一类不饱和脂肪酸,月经前PGs浓度升高是导致PD的因素之一。PGE2和PGF2α与PD的机制密切相关,它们参与了子宫平滑肌松弛和收缩的调节。研究表明,PGF2α与子宫肌层中的雌二醇水平呈正相关,雌二醇水平的升高会加速PGF2α的合成和释放■◆★,并引起子宫血管痉挛,导致PD。研究发现PD患者体内花生四烯酸的异常释放会导致子宫内白三烯和PGs的级联反应■◆◆★◆◆。这些白三烯和PGs是介导PD炎症反应的底物,会引起痉挛、恶心和其他症状。此外当二十二碳六烯酸被COX和脂氧合酶代谢时,它可能会通过抑制花生四烯酸级联反应产生抗炎作用■■■◆◆◆。15 d-PGJ2◆■◆◆、血栓素B3、亚硫酸三烯B4◆★、12-氧代-20-羧基-亮三烯B4、花生四烯酸和二十二碳六烯酸含量的显著变化◆■,表明PD大鼠存在炎症级联反应★★,DGSND可抑制COX-2和5- LOX,通过恢复上述脂肪酸来改善炎症反应■■★◆◆,从而对PD发挥治疗作用。虽然发现花生四烯酸在PD中的含量下调■◆★,这与之前的研究结果一致,但推测这可能是由于PD微环境中大量产生PGF2α等下游代谢产物,导致花生四烯酸含量减少◆■◆★,而DGSND逆转了这种消耗的减少。
图10 DGSND通过调节脂质稳态缓解PD的潜在机制★■★◆。红色字体表示DGSND上调的代谢物★■★,蓝色前词表示DGSND 下调的代谢物/酶。
本研究通过结合行为指标、生化指标■■◆、子宫指数和子宫病理指标来评价DGSND对PD的改善作用。在实验结束后,DGSND组的扭动次数和潜伏时间与模型组相反(图2A-B)★◆◆,PGF2α/PGE2值、子宫指数和病理变化也与模型组相反(图2C-E)。PGF2α/PGE2值的升高值是临床诊断PD的一个重要参数。经DGSND治疗后■■,上述指标均明显改善,并显示出与POS组相似的保护作用。
通过以下阈值筛选出潜在的生物标志物:VIP1.0,P0■■★.05。然后通过分析其保留行为、精确分子量★★★■◆、MS/MS片段信息以及METLIN()、人类代谢组数据库5.0()等各种数据库对其化学结构进行鉴定★★◆◆。利用上述代谢物鉴定方法■★★,以吲哚丙烯酰甘氨酸为例其鉴定过程如图S4所示。最后共鉴定出35种尿液代谢物,其中模型组的4-aminohippuric acid、γ-CEHC■■◆◆、雄烯二酮等19种代谢物的含量较对照组升高。同时与对照组相比,模型组中吲哚丙烯酰甘氨酸、6-羟基多巴胺■★、S-腺苷半胱氨酸等16种代谢物含量降低■◆■◆■★。表S7显示了潜在生物标志物的具体信息,图S5和图S6显示了这些生物标志物相对含量的详细变化和热图。
组胺是肥大细胞的代谢产物,可由组氨酸经组胺脱羧酶产生★★,而肥大细胞广泛分布于子宫肌层★★◆。组氨酸含量升高导致组胺含量增加,从而引起子宫肌层收缩,导致腹膜透析◆★★。本研究中★■◆★★◆,PD大鼠组氨酸含量显著升高,表明子宫平滑肌处于收缩状态,而DGSND可逆转组氨酸的异常状态。此外本研究中检测到的3-羟基邻氨基苯甲酸★■◆■、吲哚乙酸、黄原酸、N-乙酰色氨酸和色氨酸醇与色氨酸代谢密切相关◆★★■。据报道作为上述代谢物的合成前体,色氨酸与干扰素-γ呈负相关,说明色氨酸与炎症有一定的关系。研究还表明色氨酸代谢与子宫内膜细胞密切相关。酪氨酸最重要的功能之一是转化为儿茶酚胺,如肾上腺素、去甲肾上腺素和多巴胺■★◆★◆。肾上腺素可作用于子宫平滑肌受体,激活腺苷酸环化酶◆★★★◆,进而增加细胞内cAMP的浓度◆■■★◆★,导致子宫平滑肌松弛。PD大鼠尿液中肾上腺素含量明显降低◆★◆,这可能会导致子宫平滑肌收缩◆★■,进而表现出PD的症状★◆■◆★。高水平的去甲肾上腺素和多巴胺可激活催产素神经元★◆◆◆,增加催产素的释放。催产素不仅直接作用于子宫肌细胞引起收缩,还能刺激子宫内膜细胞释放PGs并导致PD。硫酸去甲肾上腺素和6-羟基多巴胺作为去甲肾上腺素和多巴胺的代谢产物,在本研究中PD大鼠尿液中含量显著降低,这可能是由于上游酶活性紊乱,导致去甲肾上腺素和多巴胺在体内蓄积★■■■■◆,从而引起PD。
Spearman相关分析了所有差异表达和极其相关的脂质(r2 0■■◆.9)■◆★★。如图8所示★★,lysoPC (P-18:0)★■■、PC(16:0/20:4)■■◆、PC(18:0/18◆◆■■:1)、PC(16■★■◆:0/20:3)和二十碳三烯酸表现出显著的相关性(edges45)◆■★★◆。不同种类的脂质之间存在显著的相关性,其中PE★■■★◆、PC和FA的相关性较高。PC和PE是哺乳动物细胞膜中含量最高的磷脂■◆■◆★■,FA也是细胞膜的结构成分◆■★◆■。虽然目前还不清楚脂质之间的关系,但lysoPC被认为是与妇科疾病有关的生物标志物。脂质之间的高度相关性表明,DGSND通过不同种类脂质之间的相互作用调节脂质平衡。
图4 DGSND干预后尿液代谢谱的多元统计分析和Ingenuity路径分析■◆。(A-B)正负离子模式下对照组■◆★◆、模型组和DGSND组PCA评分图■■■;(C)对照组、模型组和DGSND组潜在尿液生物标记物热图◆◆◆■■;(D) Ingenuity pathway分析。
结果:DGSND可有效改善PD大鼠的扭动时间■◆★■■◆、潜伏期、PGF2α/PGE2值、子宫指数和形态学变化★◆◆◆★★。PD大鼠的代谢特征主要以类固醇激素代谢、氨基酸代谢和脂质代谢紊乱为特征。功能分析显示PD的尿液生物标志物与脂质异常最相关■■◆◆★■。进一步的脂质组学分析表明◆◆◆,DGSND通过重塑脂质稳态发挥抗PD作用★■,这可能是由于不同种类脂质之间的显著相关性,尤其是磷脂酰乙醇胺、磷脂酰胆碱和脂肪酸的极高相关性。此外DGSND还能显著降低CK、PLA2、LPCAT3、COX-2和5-LOX等关键代谢酶的表达。
图5 子宫组织样本的多元统计分析及差异脂质的特征★◆。(A-B)正负离子模式下对照组和模型组的PCA评分图。(C-D)正负离子模式下对照组和模型组的OPLS-DA评分图■★★■◆★。(E) 负离子模式下磷脂酰甘油(16:0/18★◆★★◆:1)的质量碎片信息和可能的碎片途径。(F) PD大鼠子宫组织中差异脂质的类别划分■★★◆◆。
鞘磷脂是细胞膜的基本成分,也是细胞生长和平衡的重要调节剂★★◆◆■。鞘磷脂是细胞膜的基本成分,也是细胞生长和稳态的重要调节因子。痛经和痉挛性疼痛是PD的典型症状,这可能与大脑机制异常有关。据报道,鞘磷脂-神经酰胺代谢与神经痛有关。作为鞘磷脂生物合成和分解的核心分子◆■◆★■,神经酰胺在复杂的网络中发挥着核心作用。此外神经酰胺还被证明可以调节血管紧张度或血管收缩。鞘氨醇作为一种神经受体配体,与PGE2和PGF2α的结合有关。此外鞘脂还被认为是多囊卵巢综合征和子宫内膜异位症的风险因素。在本研究中模型组的鞘氨醇含量显著增加◆■★,而神经酰胺含量显著减少。经DGSND干预后二者均可恢复。
为了研究与PD脂质改变相关的通路,基于KEGG数据库进行了LIPEA通路分析。气泡图显示了前7条信号通路,包括鞘脂代谢、鞘脂信号通路、坏死性凋亡◆■■、甘油磷脂代谢、铁死亡、糖基磷脂酰肌醇(GPI)-锚生物合成和醚脂代谢(图7E)。DGSND-L主要影响鞘脂代谢■◆★◆、鞘脂信号通路、甘油磷脂代谢和坏死性凋亡(图7F)★◆■◆◆。DGSND-M可完全调控参与PD的通路(图7G),DGSND-H主要作用于鞘脂代谢◆■■◆■、甘油磷脂代谢◆★★■■、鞘脂信号通路★■、铁死亡、糖基磷脂酰肌醇(GPI)-锚生物合成和坏死性凋亡(图7H)■■■■★。
DGSND治疗后★◆,PD大鼠的脂质代谢轮廓与对照组接近★◆■◆,而与模型组相差甚远,表明DGSND能有效缓解PD(图6A)。此外热图还直观地显示了各组潜在生物标志物的相对含量(图6B)★◆◆★■■。DGSND-L组可以逆转85种脂质,其中26种脂质有显著性差异;DGSND-M组可以逆转104种脂质★■★■,其中83种脂质有显著性差异;DGSND-H组可以逆转104种脂质,其中79种脂质有显著性差异。通过DGSND对生物标志物的有效调节,缓解了PD的进展。
富集分析有助于深入了解不同亚类脂质的作用机制★■◆★◆◆。如图7A-D所示★■★,每个节点代表显著变化的脂质组(P0.05),大小反映了每个脂质组所含脂质的数量。与对照组相比■◆■◆★★,模型组中有14类脂质发生了显著变化(图7A)■★。结果显示★◆◆◆★,除了不饱和脂肪酸、不饱和神经酰胺■◆■◆、甘油酯◆■★、饱和PC下调,部分饱和神经酰胺和溶血PCs下调外■◆,其他脂类总体呈上升趋势(图7A)。经DGSND治疗后,各种脂质可出现不同程度的逆转★◆★,包括整体调控的PIs■■■◆★、PSs和不饱和溶血磷脂(图7B-D)。DGSND-L组可以逆转PIs、PSs、不饱和溶菌酶◆■★■◆◆、饱和神经酰胺、饱和溶菌酶和不饱和PCs(图7B);DGSND-M组可以逆转PIs、PSs、PEs★★◆◆、不饱和溶菌酶■■、不饱和FAs、不饱和神经酰胺、饱和神经酰胺、磷脂醚、不饱和PCs和磷脂酰甘油(图7C)★★■★■;DGSND-H组可以逆转PIs、PSs■■、PEs、不饱和溶菌酶、饱和溶菌酶、不饱和FAs■◆、不饱和神经酰胺■■、磷脂醚、不饱和PCs和磷脂酰甘油(图7D)◆◆★。DGSND-H组能逆转PIs、PSs、PEs◆◆★■★◆、不饱和溶血磷脂、饱和溶血磷脂、不饱和FAs■■◆★★◆、不饱和神经酰胺、磷脂醚■◆◆、不饱和PCs和磷脂酰甘油(图7D)。富集分析结果表明,不同剂量的DGSND可通过调节脂质,尤其是磷脂和脂肪酸来缓解PD。因此可以推测DGSND可能通过调节子宫组织的脂质平衡来改善PD。
众所周知,月经周期取决于卵巢激素浓度的周期性变化,而类固醇激素紊乱被认为是月经紊乱的一个重要因素。胆固醇可转化为孕烯醇酮,进而转化为孕酮★◆★★,孕酮可生成雌二醇的低活性形式--雌酮◆★■■,从而减少PGs的合成,进一步减轻子宫平滑肌收缩程度■◆■◆◆,缓解PD。雄烯二酮是类固醇的代谢产物◆◆★■★■,黄体酮在17-羟化酶的催化下可生成17-羟基黄体酮◆◆■★,17-羟基黄体酮可进一步转化为雄烯二酮。PD大鼠体内17β-雌二醇-3-葡萄糖醛酸■◆★★★、皮质醇21-硫酸盐、5α-四氢皮质醇、雄烯二酮和17α,21-二羟基孕烯醇酮水平升高,而皮质酮水平降低★◆◆,表明类固醇的生物合成受到干扰,从而导致PD症状。而在DGSND干预后则可得到不同程度的调节。
甘油磷脂是生物体内含量较高的一种脂质,具有多种生物学功能,而PC是含量最高的甘油磷脂之一◆◆■★。PC可通过CDP-胆碱途径从头合成◆■★■■★,其中CK是限速酶。本研究中PD大鼠子宫内CK活性显著升高,PC含量显著增加,DGSND治疗后可明显抑制PC合成的活化,改善脂质紊乱。PE的合成对线粒体和内质网的功能至关重要◆◆★★■。模型组中PE含量的升高可能反映了线粒体能量生成的增加和线粒体活性的改变,从而影响PD的发生和发展。PS是一种天然磷脂,具有多种抗炎功能■■◆★◆■,被广泛认为是止血和凝血的关键调节因子。在DGSND治疗后■◆■★■◆,PS水平趋于正常,提示PS可能是PD血液循环障碍的原因。此外PE、PC和FA之间也存在极高的相关性(图8)。磷脂★◆★■■、溶血磷脂和游离脂肪酸可通过“磷脂酰基链重构途径”相互转化,其中PLA2和LPCAT3是关键酶。甘油磷脂可水解为花生四烯酸和溶血磷脂,花生四烯酸可通过环氧化酶(COX)氧化为PGs,从而诱导月经期子宫肌收缩和血管收缩。结果发现PD大鼠PLA2的表达增加,可能的原因是子宫收缩时缺氧和缺血可激活PLA2。据报道消耗的花生四烯酸可通过LPCAT3补充,从而使花生四烯酸通过赖氨酸丙二酸酰化,经磷脂酰基链重构途径转化为PC,优先掺入膜中。在PD大鼠体内发现的LPCAT3过表达也证实了脂质紊乱的存在。此外花生四烯酸通过磷脂酰基链重构途径消耗,并合成促炎症的下游代谢产物,可能会导致PD大鼠体内花生四烯酸含量下降。作为一种多功能磷脂信使■★◆★◆■,LysoPC参与细胞迁移和增殖、炎症反应并影响血管平滑肌■◆◆★,还被认为是卵巢癌和其他妇科癌症的潜在生物标志物。LysoPC的变化趋势与已发表的数据非常吻合,并且可以通过DGSND进行有效调整■★◆★■◆。
方法★◆■:采用UPLC-Q-TOF/MS对DGSND进行表征◆■◆。用苯甲酸雌二醇和催产素诱导PD大鼠模型,通扭动次数、潜伏时间、生化指标、脏器指数和组织病理学的药效学研究来评价DGSND对PD大鼠的疗效;采用尿液代谢组学结合功能分析★★◆,分析DGSND对PD大鼠的治疗效果,确定关键通路,并以子宫组织为研究对象进一步进行脂质组学分析■■■★★,从脂质稳态的角度阐明DGSND的保护机制。最后使用western blot分析验证脂质代谢中关键代谢酶的表达◆★◆◆。
1★★.当归四逆汤(DGSND)是一种临床有效的治疗原发性痛经(PD)的配方;
当归四逆汤(DGSND)是治疗原发性痛经(PD)的经典处方,而DGSND对PD的改善作用及其机制尚不完全清楚。
PCA分析直接揭示了对照组、模型组和DGSND干预组之间代谢特征的差异(图4A-B)。从PCA评分图中可以看出,DGSND干预组比模型组更接近对照组,尤其是DGSND-M组。结果表明DGSND的干预能有效改变PD的进展,改善PD大鼠的新陈代谢。此外DGSND对潜在的生物标志物有不同程度的调节作用(图S7)★★◆◆,DGSND-L组逆转了33个生物标志物,其中18个有显著性差异;DGSND-M组逆转了34个生物标志物,其中22个有显著性差异■◆;DGSND-H组逆转了32个生物标志物,其中21个有显著性差异。组间聚类结果显示,DGSND干预组与对照组合为一组◆★◆,然后与模型组聚类,其中DGSND-M组的干预效果最接近对照组,这意味着DGSND中剂量对PD大鼠有更好的疗效(图4C)。此外代谢通路分析表明,DGSND主要通过调节类固醇激素代谢、氨基酸代谢和脂质代谢发挥保护作用(图S8)。